
National Cancer Center Japan
Loading...
Loading...
Research containing National Cancer Center Japan
Get data-driven expert analysis from the CB Insights Intelligence Unit.
CB Insights Intelligence Analysts have mentioned National Cancer Center Japan in 1 CB Insights research brief, most recently on May 23, 2025.
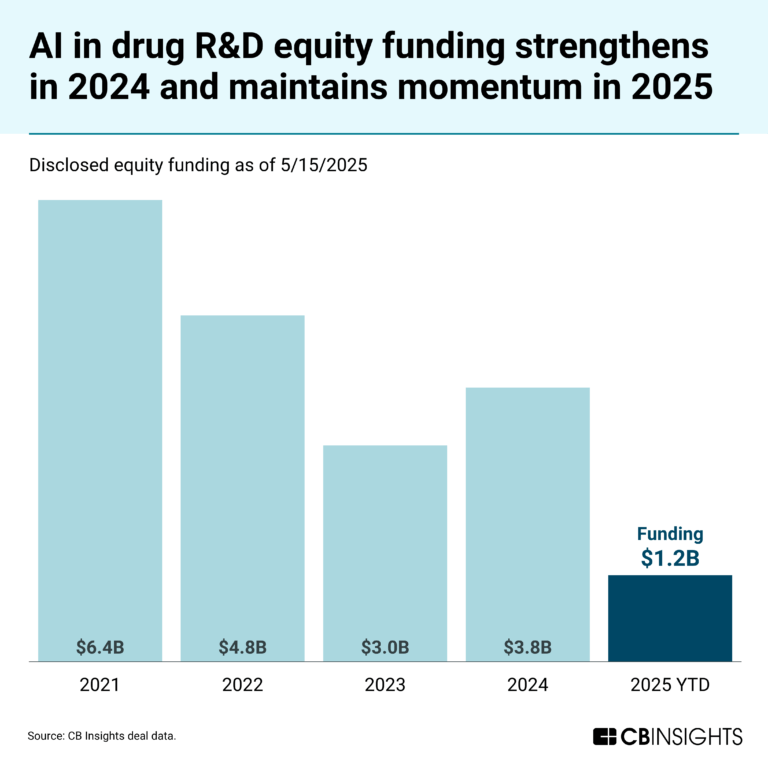
May 23, 2025
The AI in drug R&D market mapNational Cancer Center Japan Patents
National Cancer Center Japan has filed 10 patents.
The 3 most popular patent topics include:
- transcription factors
- clusters of differentiation
- experimental cancer drugs

Application Date | Grant Date | Title | Related Topics | Status |
|---|---|---|---|---|
3/7/2017 | 4/25/2023 | Experimental cancer drugs, Monoclonal antibodies, Molecular biology, Clusters of differentiation, Cancer treatments | Grant |
Application Date | 3/7/2017 |
|---|---|
Grant Date | 4/25/2023 |
Title | |
Related Topics | Experimental cancer drugs, Monoclonal antibodies, Molecular biology, Clusters of differentiation, Cancer treatments |
Status | Grant |
Latest National Cancer Center Japan News
Nov 6, 2025
Subjects Abstract RNA splicing dysregulation has emerged as a hallmark of cancer and a promising therapeutic target; however, its full landscape in human solid cancer remains poorly characterized. To address this, we perform alternative splicing analyses using RNA-sequencing data from 751 lung adenocarcinoma samples from our cohort integrated with 519 samples from The Cancer Genome Atlas. Visualization of splicing patterns using t-distributed stochastic neighbor embedding reveals substantial inter-tumor heterogeneity driven by distinct molecular subtypes and histological differentiation. We identify a unique molecular subtype associated with inactivating mutations in CMTR2, which encodes Cap-specific mRNA (nucleoside-2’-O-)-methyltransferase 2. CMTR2 mutations are observed in 3.8% of cases and are predominantly truncating mutations, which form an isolated cluster within the splicing landscape. Intrinsic and CRISPR-Cas9-engineered CMTR2 mutations disrupt alternative splicing and sensitize cancer cells to sulfonamide-based RNA splicing modulators and immune checkpoint blockade therapy. Retrospective patient data confirm the increased sensitivity of CMTR2-deficient tumors to immune checkpoint blockade therapy. These findings uncover a previously unrecognized RNA splicing deficiency in human cancers and define a molecular subtype of lung adenocarcinoma driven by RNA splicing dysregulation, suggesting targets for therapeutic intervention in lung cancer. Introduction Dysregulation of RNA splicing recently emerged as a hallmark of cancer 1 . Alternative splicing (AS), a critical process in gene expression, enables the generation of multiple mRNA and protein isoforms from a single gene, thereby supporting diverse gene functions and regulating key cellular processes. This intricate regulatory mechanism governs numerous biological functions, including tissue-specific differentiation, metabolism, neuronal development, and the pathogenesis of various diseases 2 . In malignancies, dysregulated RNA splicing drives tumorigenesis by increasing cell proliferation, inhibiting apoptosis, increasing migratory capacity, and promoting metastasis 1 . The RNA splicing process is orchestrated by the spliceosome, a large RNA-protein complex composed of small nuclear ribonucleoproteins (snRNPs) and various splicing factors. In cancer, splicing dysregulation frequently arises due to mutations in splicing factor genes 3 , 4 . In hematological malignancies, somatic mutations in genes such as SF3B1, U2AF1, and SRSF2, which are collectively referred to as “spliceosomal mutations”, are major drivers of global RNA splicing alterations 1 , 3 , 4 . Genome-wide studies have also identified spliceosomal mutations in non-hematological cancers. For example, we and others reported somatic mutations in the splicing factor genes RBM10 and U2AF1 in lung adenocarcinoma (LADC), the most prevalent histological subtype of non-small cell lung cancer (NSCLC), with mutation frequencies of approximately 7–8% and 3%, respectively 5 , 6 , 7 . Despite these discoveries, the full landscape of AS in lung cancer and the specific contributions of spliceosomal mutations remain poorly understood. In this study, we mapped the comprehensive AS landscape in LADC and explored its therapeutic implications. We analyzed RNA-sequencing (RNA-seq) datasets from 751 LADC cases from the National Cancer Center (NCC) Hospital, Japan and 519 cases from The Cancer Genome Atlas Program (TCGA, https://www.cancer.gov/ccg/research/genome-sequencing/tcga ) and visualized splicing heterogeneity across the cohorts. We performed t-distributed stochastic neighbor embedding (t-SNE) analysis of a high-dimensional percent spliced-in (PSI) value matrix for skipped exon (SE) events 8 and found distinct LADC subtypes, defined by global RNA splicing patterns. These subtypes were associated with spliceosomal mutations in RBM10 and U2AF1 and correlated with histological differentiation status. We identified a LADC subtype characterized by loss-of-function mutations in the Cap Methyltransferase 2 (CMTR2) gene. CMTR2 encodes Cap-specific mRNA (nucleoside-2’-O-)-methyltransferase 2, a key enzyme involved in methylation of mRNA and small nuclear RNA (snRNA) 9 , 10 . A previous report demonstrated that CMTR2 is significantly mutated in LADC and is enriched with loss-of-function mutations 11 . Functional analyses using both tumor-intrinsic and CRISPR-Cas9-engineered CMTR2-mutant cells showed that CMTR2 deficiency induces global alterations in RNA splicing and increases vulnerability of the splicing machinery. This heightened sensitivity renders CMTR2-deficient cells particularly susceptible to sulfonamides, which are compounds that degrade the splicing factor RNA-binding protein 39 (RBM39) 12 , 13 . We demonstrated that CMTR2-deficient LADCs exhibit increased sensitivity to immune checkpoint blockade (ICB) therapy, both in a syngeneic mouse model and in LADC patients. Results Comprehensive profiling of alternative RNA splicing in lung cancer To characterize the landscape of alternative RNA splicing events in lung cancer, we analyzed RNA-seq data from 1017 lung cancer cases in the NCC cohort using the robust rMATS computational framework (Fig. 1a ). This tool enables comprehensive identification of AS events across multiple gene sets, including SEs, alternative 5′ or 3′ splice sites, mutually exclusive exons, and retained introns (Fig. 1b ) 14 , 15 . rMATS quantifies the relative inclusion of alternative exons, splice sites, or introns and reports these as PSI values 8 , 14 . Fig. 1: CMTR2 cluster in the AS landscape of LADC. a Schematic showing the study workflow. The bar graph on the left shows the number of tumor samples of each histological type in the NCC lung cancer cohort. Abbreviations: ADC, adenocarcinoma; ASC, adenosquamous carcinoma; LCC, large cell carcinoma; LCNEC, large cell neuroendocrine carcinoma; PC, pleomorphic carcinoma; SCC, squamous cell carcinoma; SCLC, small cell lung carcinoma. b Types of AS events detected by rMATS. c–g AS landscape of LADC samples from the NCC cohort, analyzed based on SE PSI values. Each t-SNE plot reflects the reduction in the higher-dimensional splice event PSI matrix for each sample into two dimensions. t-SNE plots are color-coded according to driver genes (c), type of U2AF1 mutation (d), type of RBM10 mutation (e), smoking status (f), and type of CMTR2 mutation (g). h RNA cap structure. Methylation sites and the corresponding methyltransferases are indicated. The chemical structure was modified from CHEBI:167614 obtained from the ChEBI database ( https://www.ebi.ac.uk/chebi/ ). i Frequency of each CMTR2 mutation type in the TCGA PanCancer Atlas. Previous comparative analyses of tumor and normal tissue samples in various cancer types highlighted an increased prevalence of SE events in tumor samples, and this difference is particularly pronounced in lung cancer 16 . To visualize splicing heterogeneity in lung cancer, we applied t-SNE dimensionality reduction to the high-dimensional PSI value matrix of SE events. Histological subtypes with neuroendocrine differentiation, such as small cell lung cancer (SCLC) and large cell neuroendocrine carcinoma (LCNEC), formed closely clustered groups. Adenocarcinoma (ADC, also referred to as LADC) and squamous cell carcinoma (SCC) tended to show different distributions. Some cases showed overlap between these two histological types, possibly representing combined-type samples with each differentiation component. Additionally, pleomorphic carcinoma was scattered throughout the landscape, reflecting its nature as a histological type that contains sarcomatoid and various histological subtype components (Supplementary Fig. 1a ). These observations were corroborated by unsupervised hierarchical clustering, which revealed unique splicing patterns across different histological subtypes (Supplementary Fig. 1b ). We next focused on LADC (n = 751), the most prevalent histological subtype of lung cancer. Initial analysis of whole-exome sequencing data revealed no clear clustering of cases based on driver oncogene mutations (Fig. 1c ), suggesting that factors beyond driver oncogenes primarily contribute to the observed inter-tumor heterogeneity in AS. Somatic mutations in splicing factor genes such as RBM10 and U2AF1 have been reported in LADC 5 , 6 . In this cohort, RBM10 and U2AF1 mutations were detected in 78 (10.4%) and 8 (1.1%) cases, respectively. Cases harboring these mutations formed distinct t-SNE clusters (Fig. 1d, e ). Specifically, the U2AF1 cluster exclusively comprised cases with the S34F mutation, a hotspot mutation in the first zinc finger domain of U2AF1 5 , 6 (Fig. 1d and Supplementary Fig. 1c ). The RBM10 cluster predominantly included cases with truncating mutations, underscoring the role of RBM10 as a tumor suppressor 17 (Fig. 1e and Supplementary Fig. 1d ). The AS landscape supported the histopathological heterogeneity of LADC. For instance, a t-SNE cluster enriched for KRAS-mutant cases exhibited high expression of both HNF4A and MUC5AC, which are expressed at high levels in invasive mucinous adenocarcinoma, a distinct histological subtype of LADC 18 (Fig. 1c and Supplementary Fig. 1e, f ). This cluster frequently displayed loss-of-function mutations in NKX2-1 (Supplementary Fig. 1g ), which lead to loss of pulmonary identity and conversion to a gastrointestinal lineage 19 (Supplementary Fig. 1h ). We also examined the epithelial-mesenchymal transition (EMT), a critical histological transformation process involved in cancer progression 20 . Using an established scoring method based on the EMT gene signature 15 , 20 , we assessed EMT status across the t-SNE landscape and found that samples with mesenchymal features predominantly clustered in specific regions and had a different distribution compared with samples with epithelial features (Supplementary Fig. 2a–c ). This finding aligns with the known involvement of AS in EMT regulation 21 . A small cluster exhibited characteristics of neuroendocrine differentiation, as indicated by a high neuroendocrine score based on the neuroendocrine gene signature 22 , 23 (Supplementary Fig. 2d ). This differentiation was further characterized by increased expression of ASCL1 and concurrent upregulation of SRRM4, which encodes a splicing factor that drives neuroendocrine differentiation in prostate cancer 24 (Supplementary Fig. 2e, f ). Characterization of the alternative RNA splicing landscape revealed distinct cancer-related clusters shaped by splicing heterogeneity and associated with both spliceosomal mutations and histopathological features. Identification of CMTR2 as a putative tumor suppressor gene with smoking-associated mutations in LADC Examination of the t-SNE landscape alongside clinical data identified a distinct cluster of 15 cases in the NCC LADC cohort that was enriched with smokers (Fig. 1f ). Thirteen of these cases (86.7%) harbored mutations in the CMTR2 gene (Supplementary Fig. 3a and Supplementary Table 1 ). These mutations, particularly truncating mutations, were significantly overrepresented in this cluster (Fig. 1g ). A comparable cluster was identified in the TCGA LADC dataset (n = 519) 25 , in which 12 of 17 cases with truncating or splice-site CMTR2 mutations formed a distinct group (Supplementary Fig. 3b and Supplementary Table 2 ). CMTR2 encodes Cap Methyltransferase 2, an S-adenosyl-L-methionine (SAM)-dependent methyltransferase involved in the 5′ capping of RNA. In mRNA, the 5′ cap structure consists of N7-methylguanosine (m7G), whereas in snRNA, it consists of N2,2,7-trimethylguanosine (m2,2,7 G); both are linked to the first transcribed nucleotide via a unique 5′−5′ triphosphate bond. In vertebrates, CMTR1 and CMTR2 proteins methylate the first and second nucleotides at the 2′-O-ribose position, forming Cap1 and Cap2 RNA modifications, respectively 9 , 10 , 26 (Fig. 1h ). The Cap2 modification, which is present in approximately 50% of polyadenylated RNA in human cells, plays a crucial role in RNA stability, RNA processing, and immune evasion 27 . CMTR2 is essential for mammalian embryonic development, as demonstrated by the embryonic lethality observed in Cmtr2-knockout mice 28 , 29 . Despite its essential role in RNA metabolism, the contribution of CMTR2 to RNA splicing and tumorigenesis remains largely unexplored. To expand upon these findings, we examined TCGA PanCancer Atlas datasets 25 and identified CMTR2 mutations at notable frequencies in uterine corpus endometrial carcinoma (32/529 cases, 6.0%), LADC (32/566 cases, 5.7%), and cutaneous melanoma (25/448 cases, 5.6%) (Fig. 1i ). In cancers with relatively high CMTR2 mutation frequencies, the mutations were often associated with hypermutated subtypes characterized by POLE mutations or microsatellite instability-high (MSI-H). However, this association was not observed in lung cancer (Supplementary Fig. 3c ). To further investigate the role of CMTR2 mutations in cancer, we used the IntOGen web platform 30 , which integrates data from diverse cancer sequencing projects and employs multiple in silico methods to identify potential driver mutations. CMTR2 was identified as a putative driver gene in two independent lung cancer cohorts (TCGA LADC and the Hartwig Medical Foundation NSCLC cohort 31 ) but not in other cancer types (Supplementary Table 3 ). These results suggest that while CMTR2 mutations may be passenger mutations in hypermutated subtype cancers, they appear to have functional significance in lung cancer. Based on these findings, we reanalyzed TCGA LADC data using programs included in the IntOGen workflow. Both the MutPanning 32 and OncodriveFML 33 programs identified CMTR2 mutations as driver mutations alongside other well-established genes implicated in LADC (Supplementary Fig. 4a–c ). Clonal population analysis using PyClone-VI 34 demonstrated that CMTR2 mutations were truncal and co-occurred with key driver mutations such as those in KRAS and TP53 in the NCC LADC cohort (Supplementary Fig. 4d, e ). The proportion of truncating CMTR2 mutations, especially nonsense mutations, was significantly higher in LADC (2.5%) than in other cancer types (Fig. 1i ). These truncating mutations were frequently associated with reduced copy numbers at the CMTR2 locus, indicating loss of the wild-type (WT) allele (Supplementary Fig. 4f ). In lung cancer, stop-gain (nonsense) mutations are commonly observed in key tumor suppressor genes such as TP53 and are strongly enriched in mutational signatures linked to tobacco smoking 35 . Combined analysis of the NCC and TCGA cohorts revealed that CMTR2 mutations were significantly more frequent in smokers than in non-smokers (5.5% vs. 0.8%, respectively; Supplementary Table 4 ). Furthermore, most CMTR2 mutations were C > A transversions, consistent with a mutational signature (SBS4) strongly associated with smoking 36 (Supplementary Fig. 4g ). In summary, these findings provide insight into the role of CMTR2 mutations in LADC and confirm previous reports of CMTR2 as a putative tumor suppressor gene, adding the observation that CMTR2 mutations are mostly smoking-associated. Deleterious effects of missense mutations in CMTR2 In the NCC cohort, the t-SNE cluster harboring CMTR2 mutations included cases with missense mutations (Fig. 1g ). The CMTR2 protein has two major domains: the N-terminal catalytic Rossmann-fold methyltransferase (RFM) domain and the C-terminal non-catalytic RFM domain 37 . In the catalytic domain, the K-D-K triad motif, comprising residues K117, D235, and K275, is critical for Cap2 methylation activity 9 . Among the observed missense mutations, two variants, K117N and K275N, were located in this motif (Fig. 2a, b ). The critical role of the K-D-K triad in methylation 9 strongly suggests that these mutations disrupt the enzymatic activity of the CMTR2 protein. The L291R mutation, which was recurrent in cases within the CMTR2 t-SNE cluster, was identified as a truncal mutation (Fig. 2a and Supplementary Fig. 4e ). This finding indicates that missense mutations outside the K-D-K motif may also impair CMTR2 function. Fig. 2: Distribution of CMTR2 missense mutations within the NCC cohort. a Top: Lollipop plot showing CMTR2 mutations identified in the CMTR2 cluster in the NCC cohort. Bottom: Domain structure of the CMTR2 protein aligned with the amino acid positions shown in the lollipop plot above. b Predicted CMTR2 protein structure. The N-terminal region harboring the catalytic domain is shown in cyan and the C-terminal region harboring the non-catalytic domain is shown in green. The CMTR2 missense mutations in the CMTR2 cluster of the NCC cohort are shown in red. c Superimposition of the mean structures obtained from five 1-μs MD simulations of WT CMTR2 (green) and the K117N mutant (magenta) complexed with an mRNA substrate and SAM. The main chains of the protein and mRNA are shown as a transparent ribbon model, while the catalytic K-D-K motif residues (K/N117, D235, and K275), SAM, and the second transcribed nucleotide of the mRNA are highlighted with stick models. Black and red arrows indicate the distances between the amino nitrogen of K/N117 and the 2’-O position of the second transcribed nucleotide, respectively. d All possible missense mutations in CMTR2, with AM pathogenicity scores (y-axis) plotted against amino acid positions (x-axis). Large dots represent CMTR2 mutations in the TCGA or NCC LADC cohort; mutations in cases within and outside the CMTR2 cluster in the t-SNE plot are shown in red and blue, respectively. The bar chart above the scatter plot shows the frequency of likely pathogenic mutations at each amino acid position. e Predicted protein structure of CMTR2 with the 5′ RNA cap (green) and SAM (yellow). Left: cartoon; right: surface representation. The color of each residue represents the average AM pathogenicity score. Residues are shown in red (likely pathogenic), blue (likely benign), or gray (ambiguous). To investigate the structural impact of these mutations, we performed molecular dynamics (MD) simulations using the supercomputer Fugaku 38 and analyzed a conformational difference between the WT CMTR2 protein and the K117N or K275N mutant, focusing on a region consisting of the K-D-K triad, mRNA/snRNA 5’ cap structures, and SAM. Our simulations revealed that the geometry of an interaction network between the catalytic motif and substrate components was altered in both mutants. The mutants exhibited increased distances between the catalytic residues and the mRNA/snRNA second transcribed nucleotide, providing insight into the molecular mechanism underlying the disrupted enzymatic activity (Fig. 2c and Supplementary Fig. 5a–c ). Furthermore, to systematically evaluate the pathogenicity of all identified CMTR2 missense mutations, we used AlphaMissense (AM) to predict their functional impact 39 . Missense variants found in cases that formed t-SNE clusters in the NCC or TCGA cohort consistently exhibited high pathogenicity scores, whereas variants outside these clusters tended to exhibit lower scores (Fig. 2d, e and Supplementary Tables 1 , 2 ). High-scoring missense mutations were predominantly located in the catalytic RFM domain, although some, such as the S508P mutation in the NCC cohort, were located in the non-catalytic domain (Fig. 2a, b, d ). Truncated proteins lacking the non-catalytic domain region lack methylation function 9 . These findings suggest that the non-catalytic domains play an auxiliary but essential role in supporting the methylation activity of CMTR2. Thus, it is plausible that missense mutations in the non-catalytic domain also impair the enzymatic function of CMTR2. Taken together, these results demonstrate that missense mutations in CMTR2 have deleterious effects on the function of this enzyme, further supporting the hypothesis that CMTR2 acts as a tumor suppressor gene. RNA splicing alterations caused by CMTR2 deficiency The role of CMTR2 mutations in RNA splicing in cancer has not been investigated to date. We evaluated the impact of these mutations on AS. In both the NCC and TCGA cohorts, when normal tissue splicing data were included in the t-SNE analysis, normal and tumor tissues were clearly segregated (Fig. 3a and Supplementary Fig. 6a ). Quantification of differential splicing events between tumor clusters and normal tissues showed that clusters harboring mutations in U2AF1, RBM10, and CMTR2 had significantly more splicing alterations than WT tumors (Fig. 3b and Supplementary Fig. 6b ). Additionally, similar to the mutual exclusivity of major splicing factor mutations in solid and hematological malignancies 40 , CMTR2 mutations rarely co-occurred with RBM10 and U2AF1 mutations in both cohorts (Supplementary Fig. 7a, b and Supplementary Tables 1 , 2 ). These findings suggest that CMTR2 mutations induce splicing changes similar to those caused by spliceosomal mutations 4 . Fig. 3: RNA splicing alteration caused by CMTR2 deficiency. a t-SNE plots for LADC (tumor) samples and normal tissues from the NCC cohort analyzed according to SE PSI values. b Stacked bar charts indicating the count and type of differentially spliced events identified by comparing normal tissues with WT tumors and RBM10-, U2AF1-, and CMTR2-mutated samples (with only truncating mutations in CMTR2); n = 5 each. Data were randomly sampled from each cluster within the NCC cohort. Spliced events were defined as those with |ΔPSI | ≥ 0.1 and an FDR < 0.05. Left: known (annotated) spliced events. Right: novel (unannotated) spliced events based on known or novel splice sites. c–e Bilateral bar charts depicting the number of significant differentially spliced events between U2AF1 (c) or CMTR2 (d) mutant and WT tumors in the NCC cohort (n = 11 per group; CMTR2 mutant samples were selected from those within the CMTR2 cluster that did not harbor a combination of other splicing factor mutations) and between CMTR2-knockout and control A549 cells (e). FDR < 0.05; red: inclusion events (ΔPSI ≥ 0.1); blue: exclusion events (ΔPSI ≤ −0.1). f Volcano plot showing differentially spliced events induced by CMTR2 knockout in A549 cells. Cutoff: |ΔPSI | ≥ 0.1; FDR < 0.05. g PCA plot showing differences in AS between CMTR2-knockout clones and control cells. Each dot represents one of three replicates for each clone or control cells. PCA analysis was performed using the PSI value of each cell. In myeloid malignancies and lung cancer, spliceosomal mutations often lead to specific aberrant splicing patterns. For example, SF3B1 mutations primarily result in reduced intron retention and differential 3′ splice site usage, whereas SRSF2 and U2AF1 mutations mainly induce alternative SE events 41 , 42 , 43 . Consistent with previous reports, splicing events that were markedly altered in U2AF1-mutated cases in the NCC and TCGA cohorts were enriched for SE events (Fig. 3c and Supplementary Fig. 6c ). Similarly, SE events dominated the splicing alterations observed in CMTR2-mutated clusters in both cohorts (Fig. 3d and Supplementary Fig. 6d ). To directly test whether CMTR2 deficiency drives these splicing alterations, we generated biallelic CMTR2-knockout cell lines from two human LADC cell lines, A549 and NCI-H3122, using CRISPR-Cas9. Three independent A549 and two independent NCI-H3122 CMTR2-knockout clones were established (Supplementary Fig. 8a–d ). Additionally, CMTR2-knockout cells were generated from the mouse Lewis lung carcinoma (LLC) cell line (Supplementary Fig. 8e ). RNA-seq analysis of these clones revealed widespread splicing alterations predominantly involving SE events, mirroring the splicing changes observed in CMTR2-mutated LADC cases from the NCC and TCGA cohorts. Thousands of significantly altered SE events were observed in CMTR2-knockout cells compared with control cells (Fig. 3e, f and Supplementary Fig. 8f, g ). Principal component analysis (PCA) of SE events according to PSI values robustly separated CMTR2-knockout cells from control cells (Fig. 3g and Supplementary Fig. 8h ). These results demonstrate that CMTR2 loss-of-function mutations induce significant splicing alterations, among which SE events are the most prominent. Splicing events affected by CMTR2 deficiency Next, we examined the overlap of differentially spliced genes between CMTR2-mutated cases from the NCC and TCGA cohorts and CMTR2-knockout cell lines. Seventy-one SE events were common to both human tumor samples and knockout cell lines (Fig. 4a , Supplementary Fig. 9a , and Supplementary Data 1 ). Among these shared splicing events, SPPL2A (Signal Peptide Peptidase-Like 2 A) and SMARCA1 (SWI/SNF-Related, Matrix-Associated, Actin-Dependent Regulator of Chromatin, Subfamily A, Member 1) were selected for further validation due to their robust PCR-detectable exon length; SPPL2A showed increased exon inclusion and SMARCA1 showed increased exon exclusion (Fig. 4b and Supplementary Fig. 9b–e ). In the t-SNE AS landscape of the NCC cohort, these splicing alterations were observed specifically in CMTR2-mutated cases (Fig. 4c, d ). Furthermore, we performed long-read direct RNA-seq of A549 CMTR2-knockout cells, which demonstrated changes in read coverage consistent with short-read sequencing (Supplementary Fig. 10a–d ). Isoform quantification analysis also revealed altered ratios between exon-including and exon-skipping isoforms of these genes (Supplementary Fig. 10e–g ). These splicing changes were validated by performing RT-PCR analysis of both CMTR2-knockout cell lines and the NCI-H1373 and NCI-H1915 lung cancer cell lines, which harbor truncating CMTR2 mutations (Fig. 4e–h , Supplementary Fig. 11a–c , and Supplementary Table 5 ). Overexpression of WT CMTR2 in the NCI-H1373 cell line decreased the altered SE events (Supplementary Fig. 12a-c ). Fig. 4: Reproducible SE events induced by CMTR2 deficiency. a Venn diagram showing the overlap of significantly differential SE events in CMTR2 mutant versus WT samples in the TCGA and NCC cohorts and in CMTR2-knockout versus control A549 and NCI-H3122 cell lines. b RNA-seq read coverage plots showing differential SE events in SPPL2A (left) and SMARCA1 (right) in A549 and NCI-H3122 cells upon CMTR2 knockout. c, d t-SNE plots of LADC samples from the NCC cohort were analyzed based on SE PSI values and color-coded according to the SE PSI of SPPL2A (c) and SMARCA1 (d). e Agilent TapeStation gel-like images of RT-PCR products validating differentially spliced events in SPPL2A and SMARCA1 upon CMTR2 knockout in A549 cells. f Quantification of PSI values for SPPL2A (left) and SMARCA1 (right). n = 3 independent experiments per condition. Error bars, ± SEM. Statistical significance was determined by a one-way ANOVA followed by Dunnett’s multiple comparison test. SPPL2A: ****p < 0.0001; SMARCA1: **p = 0.0043, ****p < 0.0001. g Western blot analysis of CMTR2 expression in NSCLC cell lines (representative of three independent experiments). h Comparison of SPPL2A and SMARCA1 splicing patterns in CMTR2-expressing A549 and NCI-H3122 cells and CMTR2-deficient NCI-H1373 and NCI-H1915 cells (representative of three independent experiments). i, j Violin plots depicting PSI values for SPPL2A (i) and SMARCA1 (j) SE events across different tumor types from the TCGA cohort: LADC, lung adenocarcinoma (n = 518); LUSC, lung squamous cell carcinoma (n = 499); BRCA, breast invasive carcinoma (n = 1044); and SKCM, skin cutaneous melanoma (n = 462). Red and blue plots indicate samples harboring CMTR2 truncating mutations. Black lines indicate median values. Exon inclusion in SPPL2A and exon exclusion in SMARCA1 were observed not only in LADC, but also in MDA-MB-231 cells, a breast cancer cell line with a truncating CMTR2 mutation (Supplementary Fig. 13a, b and Supplementary Table 5 ). Increased exon inclusion in SPPL2A and increased exon exclusion in SMARCA1 were observed in multiple cancer types in the TCGA dataset (Fig. 4i, j ). These findings suggest that CMTR2 deficiency alters the splicing of diverse genes and that certain genes are commonly affected in different cellular contexts. CMTR2 interactions with splicing machinery We next investigated the mechanistic link between CMTR2 and splicing dysregulation. Analysis of protein–protein interaction databases 44 , 45 indicated there are physical interactions between CMTR2 and spliceosomal complex components, particularly snRNPs (Supplementary Tables 6 , 7 ). To validate this, we generated Flp-in T-REx 293 cells expressing FLAG-tagged WT CMTR2 or the K117N mutant, a loss-of-function variant identified in our NCC cohort, under doxycycline control (Fig. 5a, b ). FLAG immunoprecipitation was performed to isolate CMTR2 and its interacting proteins. SDS-PAGE separation of immunoprecipitates revealed protein bands specific to WT CMTR2 samples (Fig. 5c ). Mass spectrometry analysis of these bands identified enriched U1 snRNP proteins in WT CMTR2 complexes, confirming that the association of U1 snRNP with the K117N mutant was impaired (Fig. 5d and Supplementary Data 2 ). Western blotting of immunoprecipitated complexes confirmed that the U1 snRNP component SNRNP70 did not bind to the K117N mutant (Fig. 5e ). Fig. 5: Interactions between CMTR2 and spliceosomal components. a Schematic showing the experimental workflow for NanoLC-MS/MS analysis of FLAG-immunoprecipitated proteins following doxycycline (Dox)-induced expression of WT CMTR2 or K117N mutant proteins in Flp-in T-REx H293 cells. b Western blot analysis confirming Dox-induced expression of CMTR2 and FLAG proteins (representative of three independent experiments). c Silver-stained SDS-PAGE gel of FLAG immunoprecipitates indicating the bands excised for NanoLC-MS/MS analysis. Numbered bands correspond to background (1–9), WT CMTR2-specific (10–16), and K117N mutant-specific (17–18) proteins (data from a single experiment). d Gene Ontology (GO) enrichment analysis of proteins specifically enriched in FLAG-WT CMTR2 immunoprecipitates. e Western blot analysis of SNRNP70 co-immunoprecipitation with FLAG-WT CMTR2 or K117N mutant protein (representative of two independent experiments). f Schematic showing the potential interaction between CMTR2, snRNP, and pre-mRNA based on the predicted CMTR2 protein structure (UniProt ID: Q8IYT2) from the AlphaFold Database and the complex structure of snRNP (PDB ID: 7B0Y). g Bubble plot showing GSEA of transcriptome data from CMTR2-knockout clones compared with control A549 cells. GO cellular component gene sets that achieved statistical significance (FDR < 0.05) are shown. Figure panel a was created in BioRender (Nukaga, S. (2025) https://BioRender.com/jqfhjfm ). These results suggest that CMTR2 deficiency potentially affects splicing through mechanisms related to physical interactions between CMTR2 and snRNPs (Fig. 5f ). Consistently, the 71 shared SE events exhibited GT-AG splice sites characteristic of major introns targeted by U1 snRNP 1 (Supplementary Fig. 14 ). To further explore the impact of CMTR2 deficiency on the splicing machinery, we performed differential gene expression analysis followed by Gene Set Enrichment Analysis (GSEA) of CMTR2-knockout and control A549 cells. This analysis revealed that core genes involved in spliceosomal complexes, such as those encoding snRNP subunits, SR proteins, hnRNPs, the PRP19 complex, and LSm proteins, were significantly upregulated in CMTR2-knockout cells (Fig. 5g and Supplementary Fig. 15a, b ). Spliceosome-related gene enrichment was also observed in murine LLC cells, with substantial overlap of core enriched genes between species (Supplementary Fig. 15b, c ). These findings demonstrate that CMTR2 interacts with components of the splicing machinery, indicating that CMTR2 deficiency affects broad splicing networks. CMTR2 deficiency and sensitization to splicing inhibition Next, we investigated whether CMTR2 deficiency is associated with potential therapeutic vulnerabilities in lung cancer. We analyzed drug sensitivity data from the DepMap portal 46 , which includes information on a wide range of human cancer cell lines, including two NSCLC lines (NCI-H1373 and NCI-H1915) with truncating mutations in CMTR2. A search for drugs with low area under the curve (AUC) values in these cell lines identified indisulam, which displayed a notably low AUC value among 57 NSCLC cell lines (Fig. 6a and Supplementary Fig. 16a ). Indisulam is a sulfonamide-based anticancer agent that targets and degrades the splicing factor RBM39 12 . RBM39 degradation by indisulam induces splicing abnormalities by disrupting the RNA-binding protein interactome, which includes several proteins involved in mRNA splicing, and causes synthetic lethality in tumors with spliceosomal mutations 47 . Thus, our findings suggest that CMTR2 deficiency is a potential therapeutic target for RBM39 degraders. Indeed, indisulam induced apoptosis and growth suppression specifically in CMTR2-deficient cells, although it degraded RBM39 protein independently of CMTR2 status (Supplementary Fig. 16b, c ). Indisulam also increased exon skipping and intron retention in these two cell lines, which is consistent with previous reports 12 (Supplementary Fig. 16d–g ). Fig. 6: Therapeutic vulnerability of CMTR2-deficient cells to the splicing modulator indisulam. a Scatterplot showing drug sensitivity according to AUC differences (NSCLC cell lines: NCI-H1373 or NCI-H1915). b Schematic of drug sensitivity testing using patient-derived organoids (scale bar, 100 μm). c Western blot analysis of CMTR2 expression in lung cancer organoids. d Drug-response curves of CMTR2 mutant (KOR558) and WT (KOR165 and KOR391) lung cancer organoids after 7 days of treatment with indisulam (n = 4 technical replicates; mean ± SD). e–g Drug-response curves of CRISPR-mediated CMTR2-knockout clones and control cells [A549 (e), NCI-H3122 (f), and LLC (g)] after 5 days of treatment with indisulam (n = 4 technical replicates; mean ± SD). h, i Colony formation assay using CRISPR-mediated CMTR2-knockout and control A549 cells treated with indisulam for 11 days. Representative images (h) and quantification of the relative colony area for all clones (i) are shown. j Western blot analysis of CRISPR-mediated CMTR2-knockout clones and control cells (A549) after treatment with increasing doses of indisulam for 72 h (representative of three independent experiments). k Schematic showing the drug treatment and inoculation schedules. l, m Charts showing tumor volume in BALB/c-nude mice bearing control (l) or CRISPR-mediated CMTR2-knockout (m) A549 cells treated with indisulam. Day 0 represents the time of tumor cell implantation. n = 8/group. NS: not significant, two-way repeated measures ANOVA. Data are presented as the mean ± SEM. n Western blot analysis of NCI-H1373 cells transduced with CMTR2 (CMTR2-OE) or mock control and treated for 72 h with increasing doses of indisulam. o Drug-response curves of CMTR2-OE or mock control cells treated for 5 days with indisulam. p Horizontal bar chart showing the number of significant differentially spliced events in NCI-H1373, NCI-H1915, and A549 CMTR2-knockout and control cells treated with indisulam (FDR < 0.05 and |ΔPSI | ≥ 0.1). For i and o: n = 4 technical replicates, mean ± SD. The P-value was calculated using a two-way ANOVA. Asterisks indicate significant differences between the control and experimental groups at each concentration (Bonferroni’s multiple comparisons test; ****p < 0.0001, * p = 0.013). Figure panels b and k were created in BioRender (Nukaga, S. (2025) https://BioRender.com/179bqky , https://BioRender.com/ . hmgp2kd). To validate the association between CMTR2 deficiency and indisulam sensitivity, we used lung cancer organoid models 48 because tumor organoids retain the properties of the original tumors better than long-term cultured cell lines. The lung cancer organoid library contained a LADC organoid derived from a smoker that harbored a nonsense mutation (C > A transversion, AA change) in CMTR2 alongside an oncogenic mutation in EGFR (Supplementary Table 8 ). This organoid was more sensitive to indisulam than EGFR-mutant organoids without CMTR2 deficiency (Fig. 6b–d ). To confirm the link between CMTR2 deficiency and increased drug sensitivity, we evaluated drug responses in CMTR2-knockout cells. These cells showed enhanced indisulam-induced growth inhibition and increased apoptosis (Fig. 6e–j ). Additionally, in a xenograft mouse model generated by subcutaneously transplanting CMTR2-knockout A549 cells into immunodeficient mice, indisulam had favorable therapeutic effects (Fig. 6k–m ). By contrast, overexpression of WT CMTR2 in NCI-H1373 cells reduced drug sensitivity (Fig. 6n, o , and Supplementary Fig. 12a ). Similar results were obtained with a different sulfonamide compound, E7820 49 (Supplementary Fig. 17a–g ). Consistently, CMTR2-knockout A549 cells showed significantly enhanced sensitivity to two independent RBM39-targeting small interfering RNAs (siRNAs) compared with control cells (Supplementary Fig. 18a–d ). Indisulam induced global splicing alterations in both A549 control and CMTR2-knockout cells (Supplementary Fig. 19a–d ), and the number of differential splicing events was higher in CMTR2-knockout cell lines than in A549 control cells (Fig. 6p ). This suggests that CMTR2 deficiency sensitizes cells to sulfonamides by exacerbating splicing disruptions resulting from RBM39 degradation, similar to the effects observed in SF3B1-mutant cells treated with sulfonamides 47 . To further explore the vulnerability of CMTR2-deficient cells, we used gene effect scores from CRISPR knockout and RNA interference (RNAi) screens in the DepMap portal. After excluding genes with common vulnerabilities across all registered NSCLC cell lines, we identified 36 and 22 vulnerability genes from the CRISPR and RNAi screens, respectively (Supplementary Fig. 20a, b ). Enrichment analysis of these vulnerability genes revealed significant enrichment in mRNA splicing pathways and spliceosomal complex-related gene sets (Supplementary Fig. 20c, d ). Analysis of the STRING biological interaction network 44 further revealed interactions between RBM39 and the identified vulnerability genes in the mRNA splicing gene set (Supplementary Fig. 20e ). Taken together, these findings suggest that CMTR2-deficient cells show increased vulnerability in relation to their splicing machinery. Sulfonamides exert antitumor effects by targeting this vulnerability through degradation of RBM39. CMTR2 deficiency enhances responses to ICB therapy Disruptions in RNA splicing in cancer cells can generate tumor-specific neoantigens, which may elicit immune responses 16 . For example, uveal melanoma, which harbors RNA splicing disruptions due to mutations in SF3B1, generates splicing-derived neoantigens 50 . It is thus plausible that CMTR2-deficient tumors, which exhibit extensive AS alterations, could respond favorably to ICB therapy. To test this hypothesis, we utilized a syngeneic mouse model in which LLC cells were transplanted into C57BL/6 mice. Both CMTR2-knockout and WT LLC cells were subcutaneously implanted, and the therapeutic effects of indisulam and/or anti-PD-1 antibody treatment were evaluated (Fig. 7a ). Treatment with the anti-PD-1 antibody significantly suppressed growth of CMTR2-knockout LLC cells but had no effect on parental LLC cells, consistent with the reported resistance of LLC cells to anti-PD-1 antibody monotherapy 51 (Fig. 7b–e ). CMTR2-knockout LLC cells also demonstrated increased sensitivity to indisulam, further supporting the role of CMTR2 deficiency in sulfonamide responsiveness observed in our previous experiments (Fig. 6k–m ). Fig. 7: Efficacy of ICB therapy in CMTR2-deficient tumors. a Schematic showing the drug treatment and inoculation schedules. b, c Tumor volume in C57BL/6 mice bearing control (b) or CRISPR-mediated CMTR2-knockout (c) LLC cells treated with vehicle, indisulam, an anti-PD-1 antibody, or a combination of indisulam and an anti-PD-1 antibody (n = 10/group). Day 0 represents the time of tumor cell implantation. Data are presented as the mean ± SEM. d Boxplot showing the tumor volume from (b) on day 24. The center line indicates the median value, the lower and upper hinges represent the 25th and 75th percentiles, respectively, and the whiskers denote the 1.5× interquartile range. Statistical analysis was performed using the two-sided Wilcoxon rank-sum test: DMSO + isotype versus indisulam + anti-PD-1, *p = 0.019. e As in (d) but using data from (c). DMSO + isotype versus DMSO + anti-PD-1, *p = 0.011; indisulam + isotype, **p = 0.0011; indisulam + anti-PD-1, **** p = 7.6e-05; indisulam + isotype versus indisulam + anti-PD-1, *p = 0.035. f Efficacy of ICB therapy in non-squamous NSCLC cases harboring a CMTR2 truncating mutation. Datasets were obtained from Hellman et al. 52 , Anagnostou et al. 53 , and Ravi et al. 54 . Top: PFS of cases treated with an immune checkpoint inhibitor (ICI). PR partial response, SD stable disease, PD progressive disease, DCB durable clinical benefit, NDB no durable benefit, PD-L1 programmed death-ligand 1, TPS, tumor proportion score. DCB was defined as SD or PR lasting longer than 6 months; all other cases were considered to have NDB. Figure panel a was created in BioRender ( https://BioRender.com/ . w53g680). In both the NCC and TCGA cohorts, LADC cases harboring RBM10, U2AF1, or CMTR2 mutations exhibited numerous splicing abnormalities, and the frequency of abnormal splicing events was higher in CMTR2-mutant tumors than in other spliceosomal mutant tumors (Fig. 3b and Supplementary Fig. 6b ). Based on these observations, we retrospectively analyzed the efficacy of ICB therapy in NSCLC patients with truncating CMTR2 mutations. In three datasets that included both ICB therapy response data and somatic mutation profiles derived from whole-exome sequencing 52 , 53 , 54 , we identified 11 non-squamous NSCLC cases with truncating CMTR2 mutations. The disease control rate was 90.9% (10/11) in patients with truncating CMTR2 mutations compared with 68% (237/350) in those without. Durable clinical benefit was achieved in 64% (7/11) of patients with truncating CMTR2 mutations compared with 46% (171/373) of those without. Median progression-free survival (PFS) was 9.2 months for patients with truncating CMTR2 mutations compared with 5.1 months for those without (Fig. 7f ). While the difference of PFS was not statistically significant by the log-rank test due to the limited sample size (p = 0.471), the consistent trend toward improved clinical outcomes suggests that CMTR2 deficiency is associated with increased responsiveness to ICB therapy. Additionally, we identified an institutional case harboring a truncating CMTR2 mutation with available ICB treatment information, in whom there was a clinical response to pembrolizumab (an anti-PD-1 antibody) treatment (Supplementary Fig. 21 and Supplementary Data 3 ). Discussion In this study, we identified CMTR2 as a therapeutic target gene for LADC by integrating RNA splicing metrics and mutation data from our Japanese cohort and the TCGA LADC cohort. In a previous report, loss-of-function mutations in CMTR2 were observed in LADC 11 . Supporting this observation, mutations in CMTR2 were detected in 3.8% (48/1270) of LADC cases, and the majority were truncating mutations, which formed an isolated cluster within the RNA splicing landscape. The CMTR2 protein plays a crucial role in Cap2 2’-O-ribose methylation of the 5’-cap structure of RNA, a key post-transcriptional modification. CMTR2 mutations caused significant alterations in RNA splicing, thereby contributing to the formation of LADC with distinct molecular subtypes. This RNA splicing landscape offers an approach to understanding tumor heterogeneity and identifying potential therapeutic targets. This study provides evidence of a biological link between CMTR2 mutations and RNA splicing abnormalities. The involvement of CMTR2 in carcinogenesis was suggested in previous studies. For instance, loss of CMTR2 promotes proliferation of KRAS-driven lung cancer cells 55 . Germline loss-of-function variants of CMTR2 are significantly associated with an increased risk of LADC and cutaneous melanoma 56 . Additionally, the TRACERx study, a comprehensive multi-sampling analysis of lung cancer, identified CMTR2 as a target of truncal mutations 57 . Consistent with these findings, the results of this study indicate that most CMTR2 mutations are truncal, supporting the hypothesis that CMTR2 functions as a tumor suppressor gene during LADC development. To investigate how CMTR2 deficiency contributes to tumor formation, we conducted differential gene expression and pathway enrichment analyses using clinical specimens and cell models. While CMTR2-knockout cells exhibited significant changes in expression of RNA splicing-related cellular components, gene sets related to molecular functions or biological processes were not significantly enriched. These results highlight the challenges of understanding how CMTR2 deficiency affects carcinogenesis through gene expression data and suggest that alternative investigative approaches are needed. We also observed that CMTR2 mutations were more common in smokers than in non-smokers. Previous reports indicate that stop-gain mutations are enriched in a single base substitution signature associated with smoking, specifically affecting 34 tumor suppressor genes involved in cancer hallmark pathways 35 . CMTR2 was identified as one of these 34 genes. Furthermore, our analysis of TCGA PanCancer Atlas data revealed that LADC is the cancer type with the highest frequency of truncating CMTR2 mutations, further supporting the notion that CMTR2 is a target of smoking-related mutagenesis in LADC. Our immunoprecipitation experiments and protein–protein interaction database 45 analysis showed that the CMTR2 protein physically interacts with multiple components of snRNPs, which comprise snRNAs and associated proteins. U1, U2, U4, and U5 snRNAs are fully or nearly fully Cap2 methylated in different cell types 58 . Knockout of CMTR2 in HEK293T cells results in loss of Cap2 methylation on U1 snRNA 27 , whereas Cap2 modification of U2 snRNA is essential for the formation of spliceosome complexes and RNA splicing 59 . Therefore, the absence of Cap2 modification in snRNAs due to CMTR2 deficiency may destabilize the spliceosome complex and contribute to RNA splicing anomalies. CMTR2 knockout altered the expression of a broad range of splicing-related genes beyond snRNPs, indicating that CMTR2 orchestrates the global spliceosomal network. These findings uncover a previously unrecognized RNA splicing deficiency in human cancers. Our splicing analysis, conducted primarily through short-read RNA-seq, revealed characteristic splicing abnormalities associated with CMTR2 deficiency; however, short-read-based tools have inherent limitations, including potential false positives and false negatives, with respect to resolving complex splicing events such as trans-splicing, as well as quantifying accurate isoforms. Although we validated key splicing events in CMTR2-knockout cells using long-read sequencing, which showed strong concordance with the short-read data obtained from SPPL2A and SMARCA1, comprehensive genome-wide analysis of the long-read sequencing data across multiple samples was not feasible. Future systematic long-read sequencing analyses would allow more comprehensive characterization of splicing dysregulation and complexity driven by CMTR2-deficiency. Targeting RNA splicing has emerged as a promising strategy for cancer therapy. We demonstrated that CMTR2-deficient cancer cells exhibit vulnerabilities in the mRNA splicing machinery and are sensitive to RBM39 degraders, such as sulfonamides. This aligns with previous findings showing that cancer cells harboring spliceosomal gene mutations are more susceptible to pharmacological splicing perturbations 47 . Although several Phase I and II clinical trials of sulfonamides have been conducted, only mild-to-moderate anticancer effects were observed, which is likely due to the limited understanding of their pharmacologic mechanisms as RBM39 degraders and the lack of predictive biomarkers 60 . Given their high tolerability, it may be possible to demonstrate their effectiveness in clinical trials with appropriately selected patients. Sulfonamides exhibit antitumor effects through pleiotropic mechanisms 61 , and mechanisms other than RBM39 degradation might underlie their effects on CMTR2-deficient cancers. Further investigation of the mechanisms of action of sulfonamides could lead to the identification of additional therapeutic targets for CMTR2-deficient cancers. In this study, CMTR2 knockout increased sensitivity to ICB therapy in LLC cells, which are typically resistant to anti-PD-1 treatment. CMTR2-mutant tumors exhibited a higher number of both known (annotated) and novel (unannotated) splicing events. The novel splicing events generate tumor-specific RNA isoforms, which suggests that CMTR2 deficiency can result in the production of splicing-derived neoantigens. This is supported by the results of a retrospective analysis indicating that patients with CMTR2-mutant lung cancers respond favorably to ICB therapy. CMTR2 mutations could thus serve as a biomarker for identifying patients who may benefit from ICB therapy. AT1965, an inhibitor of CMTR2 that is currently in clinical trials (NCT06234098), activates B cell-mediated immune responses in a lung cancer model and shows synergistic antitumor effects when combined with PD-1 inhibition 62 . This finding further supports the notion that CMTR2 deficiency enhances the efficacy of ICB therapy. However, as a limitation of this study, the identification of neoantigens generated by CMTR2 mutations and the subsequent activation of immune responses were not experimentally validated. It is also possible that CMTR2 deficiency enhances the efficacy of ICB therapy through other mechanisms, such as aberrant splicing of immune-related genes. Additionally, we could not directly assess Cap2 methylation changes in our CMTR2-knockout models. Detailed investigation of these Cap2 alterations and their functional consequences could reveal how CMTR2 influences tumor progression and immune responses. Overall, CMTR2-deficient tumors present a therapeutic opportunity through splicing perturbation. While our preliminary data suggest that such tumors may exhibit enhanced responses to ICB therapy, larger prospective studies and mechanistic investigations are needed to validate this and to determine its clinical significance. Methods Patients The NCC cohort of 1017 cases was diagnosed with lung cancer at the NCC Hospital in Tokyo, Japan, between 2011 and 2017. The cohort comprised 568 males and 449 females. The median age was 66 years (range: 26–89 years; males: median 65 years, range 26–89; females: median 66 years, range 35–88). Sex was self-reported during patient enrollment, and confirmed by review of medical records. Gender identity was not assessed specifically in this study. Pathological diagnosis was performed according to the 7th edition of the World Health Organization classification. Non-physician healthcare professionals used questionnaires to collect information about each patient’s exposure to smoking. Never-smokers were defined as individuals who had never smoked or had smoked fewer than 100 cigarettes in their lifetime. Fresh frozen cancerous and non-cancerous lung tissues obtained through surgical resection were available from the NCC Biobank and used for molecular analyses. Tissue samples were consumed during the analytical procedures. Residual samples, which were available for some cases, are held at the NCC Biobank and can be shared with other researchers under a collaborative research framework, subject to approval from the NCC Biobank and the Institutional Review Board of the NCC. Contact the corresponding authors for sharing requests. No sex or gender analysis was carried out in this study because the research questions focused on molecular mechanisms of lung cancer that are not thought to differ according to sex or gender. Participants received no financial compensation. RNA-seq Total RNA was extracted from frozen normal lung tissues or tumor samples from the NCC cohort using an AllPrep DNA/RNA Mini Kit (QIAGEN). In addition, total RNA was extracted from cell lines using an RNA RNeasy Mini Kit (QIAGEN). Short-read sequencing Poly(A)-selected strand-specific libraries were prepared using a TruSeq Stranded mRNA library prep kit (Illumina). Sequencing was performed using the Illumina NovaSeq6000 platform, with 150-bp paired-end reads. For TCGA transcriptome data, sequencing data were obtained from the Genomic Data Commons (GDC). Alignment to hg38 (reference genome) was performed using STAR ver. 2.7.9a 63 . Gene fusions were estimated using STAR-Fusion ver. 1.11.0 64 . Read counts and expression levels, normalized to transcripts per kilobase million, were estimated using StringTie ver. 2.0.4 65 or RSEM ver. 1.3.3 66 . Long-read direct RNA-seq Sequencing libraries were prepared using a SQK-RNA004 kit (Oxford Nanopore Technologies) according to the manufacturer’s instructions. The prepared libraries were sequenced on a PromethION 2 Solo instrument (Oxford Nanopore Technologies) over 72 h. Base calling was performed using Dorado basecaller ver. 0.9.1 + c8c2c9f 67 . Low-quality reads (average Q < 7) were filtered using chopper 68 . Alignment to hg38 was performed using minimap2 ver. 2.28-r1209 69 . Transcript annotation, novel isoform discovery, and expression quantification were performed using Bambu ver. 3.8.3 70 with GENCODE release 41 comprehensive annotations as the reference. AS analysis Detection of AS events and calculation of PSI values, also known as the exon inclusion ratio, were performed using rMATS turbo ver. 4.1.2. The detailed types of AS events that can be detected using rMATS were described in the original paper 14 . rMATS in a non-standard mode, which does not require two conditions, was used to calculate PSI values for t-SNE and PCA. All of the BAM files to be processed were listed in the --b1 file, omitting the --b2 file, and used the optional argument --statoff. The default statistical comparison mode was used to detect differentially spliced events in cell lines and human samples. For dimensionality reduction analyses (t-SNE and PCA), the following filters were applied to PSI values calculated by rMATS, and only significant AS events that passed these filters were used: average PSI values between 0.05 and 0.95 to exclude constitutive splicing events, average sum read count (inclusion count and skipping count) ≥10 to ensure sufficient coverage, percentage of samples with a missing PSI value < 0.05 to minimize the impact of missing data, and delta PSI values (maximum PSI values – minimum PSI values) >0.3 to focus on events with high biological variability. These stringent criteria ensured robust and unbiased dimensionality reduction. t-SNE was performed using the R package Rtsne ver. 0.16. PCA was performed using the prcomp function in R ver. 4.2.2. Sequence logs were created with ggseqlogo R package ver. 0.2 71 to visualize nucleotide frequency and conservation at splice sites. Use of public databases for AS analysis of the SPPL2A and SMARCA1 genes PSI data for normal tissues and cell lines were obtained from VastDB 72 ( http://vastdb.crg.eu/ ). For TCGA cohort analysis, PSI data from the SplAdder project 16 , obtained from the OncoSplicing database 73 , were used. The chromosomal position of the SPPL2A and SMARCA1 SE events in each database corresponds to the event detected in the present study, as shown in the source data of Supplementary Fig. 13 . Differential gene expression and gene enrichment analysis Differential gene expression analysis was performed using R DESeq2 package ver. 1.36.0 74 . Genes with an adjusted P-value < 0.05 and a |log2-fold change | >1 were considered to be differentially expressed genes (DEGs). DEGs were analyzed using Metascape 75 ( http://metascape.org ) with default parameters for tissue/cell-specific gene signature enrichment analysis based on PaGenBase 76 . GSEA was performed using the GSEA module in GenePattern 77 . The analysis utilized the MSigDB GO Cellular Component ontology gene sets. The permutation type was set to ‘gene_set’ with 1000 permutations. Core enrichment genes from GSEA were annotated with their spliceosomal complex, class, or family associations using the Spliceosome Database 78 . EMT score calculation An established method using the two-sample Kolmogorov-Smirnov test based on EMT signature gene expression levels 15 , 20 was employed to score the EMT status of NCC samples. The scoring script and signature genes were retrieved from https://github.com/Xinglab/rmats-turbo-tutorial . Samples were classified as mesenchymal (statistic EMT score > 0 with FDR < 0.05), epithelial (statistic EMT score <0 with FDR < 0.05), or intermediate (all other samples). Neuroendocrine score calculation Neuroendocrine scores were calculated by single-sample GSEA using GSVA R package ver. 3.2.0 79 as previously described 22 . The neuroendocrine gene signature was obtained from a previous report 23 . Whole-exome sequencing DNA extraction and sequencing DNA was extracted from tumor and matched normal tissues (e.g., blood or normal lung tissue) using a QIAamp DNA Mini Kit or AllPrep DNA/RNA Mini Kit (QIAGEN). For a small subset of samples for which RNA-seq data were available but DNA of sufficient quality could not be obtained, whole-exome sequencing was not performed. Exon capture was performed using SureSelect Human All Exon exome capture kits (Agilent Technologies). Tumor and matched normal tissues were sequenced on the HiSeq2500 or NovaSeq6000 platform (Illumina) using 2 × 75 bp/2 × 100 bp or 2 × 150 bp paired-end reads, respectively. The sequencing depth was set to 150× for tumor samples and 100× for normal controls. Data analysis FASTQ data were subjected to genome mapping and variant calling using an in-house data analysis pipeline. Alignment to the reference genome (hg38) was performed using Parabricks ver. 3.1.3 (Nvidia), which implements algorithms equivalent to BWA ver. 0.7.15 80 and GATK ver. 4.1.0 81 . Duplicate reads were identified and marked, followed by realignment and recalibration of base quality scores. The mapped data were then exported in BAM format. Variant calling Somatic single-nucleotide variants (SNVs) were called using Mutect2 (GATK ver. 4.1.2.0) in tumor-normal mode 82 . Variants were filtered using FilterMutectCalls and FilterAlignmentArtifacts, and only those with PASS in the FILTER field were selected. Additionally, a panel of “normals” derived from blood was used to filter background mutations. Small insertions and deletions (InDels) were called using both Mutect2 and Strelka2 83 , with only PASS-filtered variants selected. Variant annotations Detected variants were annotated using OncoKB 84 . Oncogenic variants were defined as those annotated as “oncogenic” or “likely oncogenic” in the OncoKB database. Oncogenic SNVs were included if called by Mutect2, whereas InDel variants were included if called by Mutect2 or Strelka2. Driver genes analyzed in Fig. 1c were validated using the Integrative Genomics Viewer 85 . For U2AF1 mutations, there are issues with the GRCh38 (hg38) reference build that prevent detection of these mutations 86 and therefore variant calling was performed using Mutect2, with the hg19 genome as the reference. TCGA genome and acquisition of clinical data Genomic mutations, the tumor mutation burden, the MSIsensor score, and copy number alteration data for TCGA cohorts were obtained from cBioPortal ( https://www.cbioportal.org/ ). For the TCGA LADC cohort, information about smoking history was retrieved using the R package TCGAmutations ver. 0.3.0. Computational identification of driver mutations To identify significantly mutated genes in the TCGA LADC cohort, a consensus informatics approach was performed using MutPanning ver. 2.0 32 , OncodriveFML ver. 2.1.3 33 , DriverPower ver. 1.0.2 87 , and OncodriveCLUSTL ver. 1.1.3 88 . The details of each algorithm are described in the original papers. Briefly, MutPanning and OncodriveCLUSTL are designed to detect cancer driver genes by modeling the mutation probability of each genomic position according to its neighboring nucleotide context and background mutation rate. OncodriveFML and DriverPower are designed to detect cancer driver mutations by combining functional scoring schemes with a background mutation model. The input MAF files were obtained from the GDC data portal ( https://portal.gdc.cancer.gov ), and each analysis was performed using default settings. Analysis of intratumor heterogeneity Intratumor heterogeneity in samples with CMTR2 SNV mutations from the CMTR2 cluster within the NCC cohort was analyzed using PyClone-Ⅵ ver. 0.1.1 34 with default parameters. This analysis utilized SNVs called by Mutect2 (GATK ver. 4.1.2.0) 82 from whole-exome sequencing data, along with copy number variant profiles and purity estimates calculated using Facets ver. 0.6.2 89 . Samples lacking determinable purity were excluded. To visualize subclonal evolutionary relationships, phylogenetic trees were constructed using ClonEvol ver. 0.99.11 90 . Heat-map and hierarchical clustering To visualize the diversity of exon-skipping events in the clusters identified in the NCC cohort, a heat-map with unsupervised hierarchical clustering (Spearman’s correlation distance and Ward clustering) was drawn using exon-skipping events detected by rMATS analysis. Analysis was performed using ComplexHeatmap ver. 2.12.1 in the R package 91 . Prediction of CMTR2 protein structure and mutation-induced pathogenicity The predicted structure of the CMTR2 protein (UniProt ID: Q8IYT2; CMTR2_HUMAN) was obtained from the AlphaFold Protein Structure Database 92 ( https://alphafold.ebi.ac.uk/ ). This structure was aligned with the theoretical model of the catalytic domain of CMTR2 37 to display the 5′ cap (m7GpppGGAA) on RNA and SAM. AM pathogenicity scores for all possible missense mutations in CMTR2 were obtained from the AM dataset 39 . Mutations were classified according to previously reported cutoff values: likely pathogenic (score >0.564), likely benign (score <0.34), or ambiguous (0.34≤ score ≤0.564). These scores were used to color-code the protein structure and visualize the distribution of scores across all possible missense mutations. Analysis of publicly available gene dependency and drug sensitivity data Gene effect (dependency) scores, which were evaluated by RNAi and CRISPR knockout, were obtained from the DepMap portal ( https://depmap.org/portal/ ). As defined by DepMap, a score of 0 defines a gene that is not essential, whereas a score of −1 corresponds to the median of all common essential genes. CRISPR screening (DepMap Public 22Q+Score, Chronos) and RNAi screening (Achilles+DRIVE+Marcotte, DEMETER2) data were used for gene dependency analysis. AUC values indicating drug sensitivity were calculated from the PRISM Repurposing Secondary Screen 46 . Cell lines A549 (CCL-185), NCI-H1915 (CRL-5904), NCI-H1373 (CRL-5866) and LLC (CRL-1642) cells were purchased from the American Type Culture Collection. NCI-H3122 was provided by Dr William Pao (Vanderbilt University, Nashville, TN, USA). Flp-in T-REx 293 cells (R78007) were purchased from Thermo Fisher Scientific. A549 (male), NCI-H3122 (male), NCI-H1915 (female), NCI-H1373 (male), and Flp-in T-REx 293 (female) are human-derived cell lines. LLC cells are mouse-derived. A549, NCI-H3122, NCI-H1915, and NCI-H1373 cells were cultured in RPMI-1640 medium (FUJIFILM Wako Pure Chemical), whereas LLC and Flp-in T-REx 293 cells were cultured in DMEM (FUJIFILM Wako Pure Chemical). All media were supplemented with 10% fetal bovine serum (Thermo Fisher Scientific). Cultures were maintained at 37 °C in a humidified 5% CO2 incubator. Patient-derived lung cancer organoids EGFR mutation-positive organoids with and without CMTR2 mutations were used for experiments; the organoids were retrieved from the in-house lung cancer organoid library 48 . Clinical samples used to establish organoids were obtained from patients at Keio University Hospital or Kawasaki Municipal Hospital. Organoid culture was performed based on a previous report 48 . Organoids were embedded in Matrigel (Corning) and cultured in Advanced DMEM/F12 supplemented with penicillin/streptomycin, 10 mM HEPES, 2 mM GlutaMAX, 1× B27 (Thermo Fisher Scientific), 10 nM gastrin I (Sigma-Aldrich), and 1 mM N-acetyl-L-cysteine (Sigma-Aldrich). For organoid maintenance, the medium was supplemented with the following factors and inhibitors: 50 ng/mL murine recombinant EGF (Thermo Fisher Scientific), 100 ng/mL human recombinant IGF-1 (BioLegend), 50 ng/mL human recombinant FGF-2 (PeproTech), 1 μg/mL human recombinant R-spondin1 (R&D Systems), 5% Afamin-Wnt-3A serum-free conditioned medium 93 , 100 ng/mL murine recombinant noggin (PeproTech), 500 nM A83-01 (Tocris Bioscience), and 10 μM Y-27
National Cancer Center Japan Frequently Asked Questions (FAQ)
Where is National Cancer Center Japan's headquarters?
National Cancer Center Japan's headquarters is located at 5-1-1- Tsukiji, Tokyo.
Who are National Cancer Center Japan's competitors?
Competitors of National Cancer Center Japan include The University of Tokyo Hospital.
Loading...
Compare National Cancer Center Japan to Competitors

Massachusetts General Hospital integrates patient care, medical research, and education. The hospital offers diagnostic and treatment options for various medical conditions and is involved in medical research and the training of healthcare professionals. Massachusetts General Hospital serves individuals seeking medical care, as well as those involved in or supporting medical research and education. It was founded in 1811 and is based in Boston, Massachusetts.
Kyushu University Hospital is a national university hospital that provides healthcare services for the Fukuoka City region.
Hokkaido University operates as a university with an emphasis on research across various fields, including natural sciences, social sciences, and humanities. The university offers undergraduate and graduate programs, along with research opportunities, and has campuses that are noted for their environment and education, and research philosophies like 'Frontier Spirit' and 'Global Perspectives'. It is based in Hokkaido, Japan.
Tohoku University is based in Sendai, Japan.
St. Luke's Hospital is a healthcare provider that offers services in cancer care, cardiovascular medicine, neurology and neurosurgery, orthopedics, and women's health services, along with primary care, urgent care, imaging, lab services, and therapy. St. Luke's serves the healthcare needs of the greater St. Louis area. It was founded in 1866 and is based in Saint Louis, Missouri.
Juntendo University is a private university. It provides undergraduate, graduate, and short-term programs in medicine, health and sports science, nursing, health science, liberal arts, and more. It was founded in 1838 and is based in Tokyo, Japan.
Loading...